作用机制:通过将含有编码抗原蛋白的mRNA导入人体,可以跳过复制、转录过程,直接进行翻译,形成相应的抗原蛋白,从而诱导机体产生特异性免疫应答,达到预防免疫的作用。
刺突糖蛋白(S蛋白)是新冠病毒入侵人体的关键蛋白,这种蛋白可以识别人类呼吸道上皮细胞上的ACE2蛋白,并与之相互作用,从而打开新型冠状病毒入侵人体的大门。简而言之:ACE2是新冠病毒S蛋白的受体。
新型冠状病毒mRNA疫苗是将编码新冠病毒刺突糖蛋白(S蛋白,抗原蛋白)的mRNA直接注射进入体内,能够在人体内合成S蛋白,通过模拟病毒感染刺激机体产生抗体和记忆细胞。
目前应用的mRNA疫苗主要有两种形式:
①基于mRNA的传统mRNA疫苗(非复制型mRNA疫苗)
定义
:是在体外转录好的一段编码抗原蛋白的完整mRNA,上游和下游分别包含5′帽子结构和 3′poly(A)尾,只编码目标抗原。
缺点
:在体内半衰期短、抗原表达量低,需要较高的量才能诱发有效的免疫应答。
②自扩增型(self-amplifying, SAM)mRNA疫苗(又称复制子)
定义
:是通过基因工程改造的 mRNA病毒基因组,其中编码RNA复制机制的基因是完整的,用编码抗原蛋白的mRNA代替了原病毒的结构蛋白编码基因,可以在体内实现自我扩增,很少的量就可以诱发有效的免疫应答。
【应用】:
一是RNA病毒感染性疾病,mRNA疫苗可用于传染性强、危害性大的RNA病毒感染性疾病的预防,如新冠病毒感染,目前已经在预防新冠病毒感染方面起到重要作用。
二是DNA病毒感染性疾病,mRNA疫苗可用于巨细胞病毒等DNA病毒感染性疾病的预防。
三是mRNA疫苗还可用于链球菌和结核杆菌等细菌感染性疾病。
【优点】:
②外源mRNA可激活模式识别受体(pattern recognition receptor,PRR),刺激非特异性免疫反应,进而激发T、B 细胞免疫反应,具有“自我佐剂(self-adjuvant)”的特点,表现更强的免疫原性;
③与质粒DNA 疫苗相比不进入细胞核内部,只在细胞质内表达抗原,不存在整合人体基因上的风险;
④不依赖细胞培养技术,可快速构建疫苗,外源因子传播风险较低。
③裸mRNA 无法被有效递呈至细胞内并成功从溶酶体逃逸,体内体液和细胞免疫较低,因此需要脂质体增强细胞摄取并改善向细胞质转译机制的传递,从而提高mRNA 疫苗的作用。
1. mRNA进入体内后易被核糖核酸酶降解,递送技术相对复杂;
1. 有研究显示,mRNA 可激活Ⅰ型干扰素介导的先天性免疫反应,引起真核翻译起始因子2(eukaryotic initiation factor 2,eiF2)的磷酸化,导致蛋白翻译减缓并最终抑制。
2. 研究表明,一种用于预防狂犬病的mRNA 疫苗和一种编码H7N9 或H10N8 病毒血凝素的mRNA 流感疫苗在人体中的中和抗体滴度低于动物模型,有效性与动物模型差异较大。
3. 安全性方面,临床试验结果显示多个mRNA 疫苗可引起受试者轻至中度局部或全身反应,因此需要在合适的动物模型上评估mRNA 疫苗的分布,所编码的抗原在远端器官的表达及潜在的安全风险,包括对局部和全身的影响。产品安全性风险可能来源于mRNA、脂质递送系统两个方面,非临床研究中需关注脂质的毒性及在人体内诱导自身反应性抗体等。
mRNA疫苗具有高效、安全、成本较低等优点,但在身体中的表达不够稳定,可能会引起一些局部反应或全身反应等副作用,如果身体处于感冒或者发热等情况下,是不可以注射疫苗的,否则会产生不良反应。
1961年,发现mRNA
1989年,将合成mRNA包裹在阳离子脂质(带正电的脂质结构)中递送到人体细胞、青蛙胚胎
1993年,测试首个mRNA疫苗,用于预防小鼠流感
1997年,首个mRNA公司成立,Merix Biosciences,后更名为Argos,现在为Colmmune)
2001年,首次报道含四种成分的脂质纳米粒(当时用于递送DNA)
2005年,发现修饰的RNA能逃逸免疫检测,发现量产脂质纳米粒的技术
2008年,BioNTech公司成立,诺华和Shire成立mRNA部门
2012年,小鼠中测试首个脂质纳米粒包裹的mRNA疫苗
2013年,首次对预防传染病(狂犬病)的mRNA疫苗开展临床试验
2015年,首次对脂质纳米粒包裹的mRNA疫苗(流感)开展临床试验
2018年,首个使用脂质纳米粒的药物(patisiran)获批
数据来源:自U. Şahin et al. Nature Rev. Drug Discov. 13, 759–780 (2014)和X. Hou et al. Nature Rev. Mater. https://doi.org/gmjsn5 (2021).
mRNA疫苗和灭活疫苗是临床常见的两种疫苗,对于临床疾病预防意义重大。
一是制作工艺有区别,mRNA是DNA翻译成蛋白质的中间产物,mRNA疫苗是用各类病原微生物的mRNA制作,而灭活疫苗是经化学或物理等方法灭活的病原微生物制成的疫苗;
二是作用原理有区别,mRNA疫苗是一种模拟天然mRNA结构与功能,将感兴趣基因导入体内,通过诱导机体免疫应答,实现预防传染病或抗肿瘤等目的的重组疫苗,而灭活疫苗虽然在机体内失去繁殖能力,但能刺激机体产生良好免疫反应,如产生特异的抗体或致敏T淋巴细胞,进而达到预防疾病的作用;
三是效果评价有区别,病毒灭活疫苗工艺简单,但是免疫效价相对较低,mRNA疫苗制作速度快,但容易降解。
根据疫苗生产技术和工艺不同,大致可以将疫苗分为三代。
第一代疫苗:有减毒活疫苗和灭活疫苗
,减毒活疫苗是通过自然选取或人工培育致病力低下或不致病的病毒株,使原本致病的病毒或细菌毒力下降到不能致病,但可刺激人体产生免疫应答的状态,以获得对特定疾病的免疫力,常见的减毒活疫苗包括卡介苗、脊灰减毒活疫苗等,其特点是免疫效果好、作用时间长,缺点是可能存在“毒力返祖”的现象,并且对储存和运输要求较高,需要全程冷链运输保存;灭活疫苗又被称为“死疫苗”,没有致病力,可让人产生相应的疾病免疫力,常见的灭活疫苗包括百白破疫苗、新冠灭活疫苗等,其特点是安全性较好,不会出现“毒力返祖”现象,对储存和运输条件的要求不高,节省了疫苗成本,缺点是它引起机体产生的免疫力持续时间较短,通常需要多剂次接种。
第二代疫苗:是亚单位疫苗和重组基因疫苗
,亚单位疫苗是通过蛋白质水解,提取、筛选具有免疫活性蛋白质片段制成疫苗,亚单位疫苗相较全病毒疫苗来说安全性和稳定性都更好,缺点是降低了疫苗的免疫原性,可能发生抗原变性导致产生的抗体失效;重组基因疫苗是将病毒内能够诱导免疫反应发生的基因片段植入到工具细胞中,规模化的生产我们所需要的蛋白质,再收集、纯化这些蛋白质后就可以制成重组基因疫苗,如乙肝疫苗等。
第三代疫苗:是重组病毒载体疫苗和核酸疫苗
,重组病毒载体疫苗是将病毒中能够诱导合成具有免疫原型的蛋白质或亚单位的基因片段嫁接到没有致病性或致病性极低的病毒中,该载体病毒携基因片段进入人体,就能够诱导免疫反应的发生;核酸疫苗又称基因疫苗,包括DNA疫苗和mRNA疫苗,核酸疫苗是将决定病毒或细菌免疫原性的遗传物质直接导入人体,依靠人体自身细胞来产生抗原,并激活免疫系统产生免疫应答,两者的区别是DNA是先转录成mRNA再进行抗原蛋白质的合成,mRNA则是直接合成,目前在研的新冠疫苗大多属于mRNA疫苗。mRNA疫苗作为目前最新的疫苗生产技术,优势明显,不用依赖细胞扩增,制备工艺简单、研发周期短,疫苗产能得到了大大的提升,缺点是它由于mRNA极易降解,需借助特殊载体送达体内,并且对运输和储存要求也非常严格。
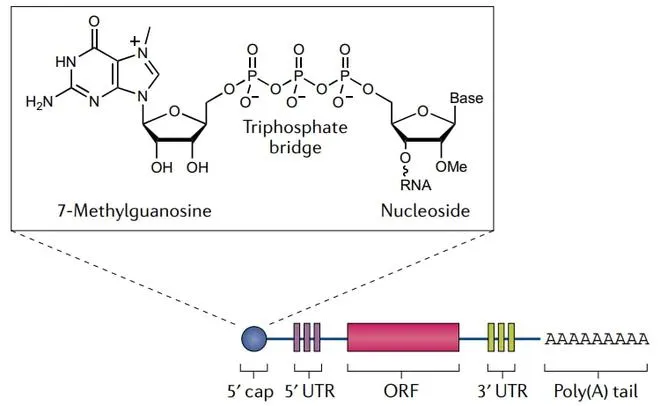
【帽子结构】
是真核生物在转录后修饰过程产生的位于5’端的特殊结构。5’端帽包含一个通过三磷酸桥连接到mRNA 5’端的7-甲基鸟苷。帽子结构包裹住mRNA的5’端,使其免于核酸外切酶的降解;还可促使mRAN环化,并募集核糖体启动翻译进程。
【非翻译区(UTR)】
包括5’UTR和3’UTR两个部分,可调节mRNA的翻译和半衰期等。5’UTR在帽子结构和编码区ORF之间,其长度和二级结构都会对翻译起始阶段的效率产生影响。3’UTR位于编码区ORF下游,在终止密码子之前。真核生物mRNA 的 3’UTR 是其重要功能元件,在翻译调控和mRNA 定位等方面起着不可忽视的作用。
【编码区ORF】
由起始密码子开始,到终止密码子结束。每三个碱基构成一个密码子,编码一个氨基酸。编码区若存在过多的二级结构和稀有密码子则会降低翻译效率。因此,“mRNA三巨头”之一的CureVac将ORF中第三个位置的A或U替换为更常见的G或C,并将这一优化策略用于其COVID-19候选疫苗CVnCoV中。但有些时候,翻译较慢的稀有密码子对保证蛋白质准确折叠很有必要,因此使用这种替换方法还须谨慎。
【3’UTR尾部】
是一段Poly A序列。与帽子结构类似,Poly(A)尾也能起到保护mRNA防止被核酸外切酶降解的作用。尾部结构也参与到翻译及其调控过程中,可与多聚腺苷酸结合蛋白(PABP)结合形成环状复合物,参与翻译的起始过程。
1. Katalin Karikó等科学家发现,在mRNA中掺入修饰核苷(假尿苷、N6-甲基腺苷、5-甲基胞苷)后可大大提高mRNA的翻译量。
2. 修饰核苷对在抑制RNA降解、提高翻译效率和调节半衰期等方面也发挥着重要作用。
3. 未修饰的mRNA(如病毒RNA)会被模式识别受体识别,从而产生免疫刺激。
辉瑞/BioNTech的mRNA疫苗BNT162b2和Moderna的mRNA-1273都利用了核苷修饰技术,用假尿苷替换尿苷。
CureVac的mRNA疫苗CVnCoV使用未经修饰的天然mRNA,通过其RNActive 平台改变mRNA序列并优化密码子,以提高mRNA的稳定性和免疫原性。
mRNA疫苗的生产可分为三大阶段,一是DNA原液制备,二是mRNA原液的制备,三是利用脂质微粒进行包封。
以辉瑞/BioNTech新冠疫苗BNT162b2的生产流程为例:
原液制备开始于质粒构建。通常使用的DNA质粒为环状质粒,质粒上含有设计好的序列模块,其中包括刺突蛋白编码。利用电流打破细胞膜,并将环状DNA质粒引入大肠杆菌。此后,大肠杆菌被储藏于含有大量营养物质的溶液中进行繁殖扩增。大肠杆菌每20分钟分裂一次,4天内可以得到数万亿DNA质粒。提取并纯化DNA质粒,过滤溶液,去除细菌及其他物质。利用酶将纯化后的环状DNA质粒切割为链状。将所得溶液分装冷藏,通过质量控制环节,并运送至下一阶段的生产加工场所。此阶段生产过程耗时10天,若将质控和运输环节纳入考虑则共耗时约17天。
目的是将DNA链转化为mRNA。上一步制备得到的DNA链与酶和核苷酸混合在10加仑容量的容器中,RNA聚合酶(RNA polymerase)会将DNA转录为mRNA。这一步骤被称为体外转录(IVT)。得到mRNA后,DNA以及其他物质将被滤除,mRNA被装进购物袋大小的塑料包装中,每袋含有约500万到1000万剂次的mRNA原料。经过经过质量控制和运输到达下一生产环节,这一阶段的生产耗时约4天,质量控制和运输耗时12天,共16天。
目的是将mRNA包裹进脂质载体(LNP)中。脂质悬浮于酒精溶液中,与mRNA接触并将其包裹,两种物质通过相反电荷相吸引。之后,原液经过切向流过滤(TFF)滤除溶液中多余的脂质、酒精等杂质,并制成最终的mRNA疫苗溶液。此生产过程耗时约12天。
辉瑞在一家位于密歇根的工厂进行这一阶段的生产,生产场地面积仅相当于一个能容纳一辆汽车的车库大小。场地内有100台混液装置工作,在30小时内制造了300万剂次的疫苗原液。
此阶段是mRNA疫苗生产的最大瓶颈之一,其中一个重要原因是市场上提供脂质的厂商有限。
在上述三个生产阶段都完成后,mRNA疫苗原液已完成,只待灌装分发。
辉瑞一管容器中包含6剂次的疫苗原液量。辉瑞位于密歇根州的工厂能够在2天内完成100万剂次的灌装。随后,经过2周的纯净度检测及其他安全性检测,疫苗便能销往世界各地。此过程共花费约19天。
1. mRNA序列结构决定抗原蛋白结构、免疫原性及稳定性
疫苗产生的抗原蛋白的序列以及稳定性决定了其激活的特异免疫的精确性和活性。而抗原蛋白的序列和结构则由mRNA序列影响和控制。因此,mRNA序列决定了疫苗的质量,也是mRNA疫苗厂商的核心竞争力之一。
生物学“中心法则”:DNA转录为mRNA转译为蛋白质。事实上,mRNA中只有部分片段会成为组成蛋白质的序列,其余部分则控制调节蛋白质的转译效率和结构。mRNA序列中,最终被转译成为蛋白质组成序列的部分被称为编码区域(translatedregion),编码区域前后分别有一个非转译区(untranslated region,UTR)。
编码区域决定了蛋白质中的氨基酸序列。编码区域中三个碱基为一组,称为密码子(codon)。密码子经转译成为特定的氨基酸,氨基酸串联后形成肽链,结构化后成为蛋白质。部分密码子组合会转译成同一种氨基酸(同义密码子Synonymouscodon),但在不同生物族群中,会存在对某一密码子组的偏好,此密码子组合的免疫原性较小,不易遭到酶类的攻击,而它的同义密码子则可能引起过敏反应并且导致质量下降。例如CAA、CAG均对应谷氨酰胺(Gln),但人类基因组中使用CAG更频繁,频率达到73%。因此CAA相较而言更有可能触发免疫反应。因此,mRNA疫苗需要选择最接近人源性的同义密码子,规避可能引起过敏反应的组合以保证安全性和转译质量。非转译区则调控mRNA和蛋白质的稳定性以及表达效率。
mRNA序列是疫苗研发中的重点,也是行业内竞争的核心。除了含有抗原蛋白编码外,序列其他部分也直接影响mRNA疫苗的质量。
编码前后的非转译区(UTR)负责调控转译以及蛋白表达,对mRNA的转译效率、半衰期、最高表达水平等数值有影响。转译效率可用来推断相同单位mRNA疫苗产生抗原的速度、半衰期用以推断mRNA能够产生抗原蛋白的时间、表达累计最高水平用以推断每单位mRNA在某一时刻能产生的抗原量的最高水平。同时,UTR中的GC水平、U水平均会影响mRNA的免疫原性,对疫苗的安全性和能否正常产生抗原造成影响。UTR需要在DNA质粒建立时包含在序列中,属于序列设计的一部分。
3’端多聚A尾(poly-A tail;A指腺苷,4种基础核苷酸的一种)位于mRNA尾部3’端,长度约100-250单位。多聚A尾能够提高mRNA的稳定性和转译效率。腺苷(A)能够降低核糖核酸酶(RNases)的效率,以此减缓mRNA的降解速度。多聚A尾可在建立DNA质粒时直接包含于编码中,也可在DNA转录为mRNA后通过聚合酶(poly-A polymerase,PAP)添加于mRNA尾部。
同时,位于mRNA最前端的5’帽结构对于降低mRNA免疫原性,增强稳定性和翻译效率有正面影响。5’帽结构是一个位于mRNA序列之前的反向7-甲基鸟苷(m7G)。在体外转录(IVT)过程中,mRNA5’端会含有三磷酸盐部分,具有很强的免疫原性。若不去除这一部分,当mRNA被递送进入人体细胞后,mRNA会在细胞质中引起IFN-1免疫反应,无法在人体内正常转译产生蛋白抗原。加帽过程可在DNA转录为mRNA的生产过程中或过程后进行,利用抗反向帽类似物(ARCA)将5’帽结构按正确方向固定在mRNA的5’端。但是此过程不能保证三磷酸盐完全被去除,所以仍旧存在引起细胞内免疫反应的风险,因此加帽工艺对疫苗的安全性有直接影响。
作为mRNA疫苗的传递介质的一大难点是mRNA以内含体的形式进入细胞质后,需要打破内含体包膜,释放mRNA。目前LNP供应商较少,且其中专利纠纷因此成为mRNA新冠疫苗快速放量的一大掣肘。
目前辉瑞/BioNTech以及Moderna所使用的LNP专利都来源于一家名为Arbutus的加拿大生物公司。Arbutus将其部分LNP专利租借给另外两家公司Acuitas和Genevant,而这两家公司分别为Moderna和BioNTech的LNP专利来源。
LNP专利成为mRNA疫苗行业商业权利的主要纷争来源!
在其他载体均有明显劣势的情况下,LNP是mRNA疫苗递送的最佳选择之一。因此,各厂商对LNP技术的掌握以及是否拥有相关专利成为了业内竞争中极其重要的一环。厂商若无相关专利,则在竞争中会处于被“卡脖子”的尴尬境地。因此,LNP技术专利决定了mRNA疫苗企业在业内竞争中的地位。
【CureVac】
CureVac公司开发了一系列基于mRNA的药物,成为mRNA肿瘤疫苗领域发展最早且全球首家建立符合GMP标准的mRNA生产线的公司。
主要研发管线包括肿瘤免疫疗法(癌症疫苗)、治疗罕见疾病的抗体疗法和针对感染性疾病的预防性疫苗。
2020年2月份,CureVac公司就宣布获得了流行病防范创新联盟 (CEPI)的支持,合作开发一种针对新冠病毒的疫苗。
2020年8月,CureVac成功在纳斯达克上市,由于拥有领先的mRNA技术,目前CureVac股价为101.73美元/股,市值达到181.05亿美元。
个性化mRNA技术涵盖了癌症免疫疗法、传染病疫苗、蛋白质替代等三个治疗平台,其中传染病疫苗包括针对新冠病毒的mRNA疫苗。
2020年3月初,辉瑞与德国mRNA技术领先企业BioNTech就开发针对新型冠状病毒疫苗的合作进行讨论。之后,双方以7.48亿美元正式达成开发协议,利用BioNTech的专有mRNA平台开发新冠候选疫苗项目BNT162。
目前,BioNTech的新冠mRNA疫苗BNT162b2已经获得美国、加拿大、欧盟、英国、以及其他多个国家/地区监管部门的批准。
2020年3月13日,复星医药和BioNTech签署合作协议,复星医药成为BioNTech在中国的战略合作伙伴,共同在中国大陆及港澳台地区开发、商业化基于其专有的mRNA 技术平台研发的、针对新型冠状病毒的疫苗产品。2019年10月,BioNTech成功登陆纳斯达克上市,目前市值达233.47亿美元。
成立于2010年,是“mRNA三巨头”中成立最晚的一个。
致力于开发mRNA药物,其候选产品跨越多个治疗领域,包括mRNA新冠疫苗,目前已经获得美国FDA的紧急使用授权。
Moderna于2018年纳斯达克上市,创造了有史以来最大生物技术IPO的记录,估值约76亿美元。到2020年,Moderna的市值已经达到488.27亿美元。
创始人是李航文博士,成立于2016年,是国内较早开展mRNA药物研发生产的企业。
自成立以来斯微生物已经完成多轮融资,尤其是去年上半年,先是获得了包括君实生物在内的3000万元A+轮融资,然后又获得了西藏药业3.51亿元的战略投资。
特点:mRNA合成平台和脂质多聚物纳米载体技术平台(LPP/mRNA)。
目前该公司有多项在研产品,包括mRNA个体化癌症疫苗、mRNA传染病疫苗、蛋白缺陷类疾病mRNA药物和遗传病mRNA药物等。
创始人是英博,成立于2019年,专注于mRNA 疫苗研发、分子设计、递送系统等技术领域,创始人曾在Moderna从事mRNA疫苗研发工作。
在融资上,艾博生物目前已完4轮融资,尤其是2020年4月由人保资本、国投创业、云锋基金共同领投的6亿元B轮融资。
特点
:该公司是目前国内唯一具有mRNA药物工业化产业化和美国FDA申报经验的团队。
在产品管线上,该公司联手军事医学科学院、沃森生物开发的新冠mRNA疫苗是国内首个获批开展临床试验的mRNA疫苗,目前该疫苗已进入II期临床尾声,将在5月与泰格医药合作开展海外III期临床试验。从2021年第三季度开始全年有1.2亿支产能,对外出口多人份产能可达到数亿剂。
成立于2019年4月,专注于mRNA药物早期创新研究,其
一步式自动mRNA药物开发平台RNApeutics平台
使蓝鹊生物在生产规模扩大和GMP质量保证方面具有一定优势。
目前蓝鹊生物与复旦大学生命科学学院和附属中山医院林金钟团队联合上海交通大学徐颖洁团队对新冠病毒mRNA疫苗展开研究,项目处于临床前阶段。
成立于2013年,最初以外包服务起家,自2018年始专注于mRNA肿瘤疫苗开发,主攻胰腺癌、直肠癌、肝癌等领域。
特点
:从肿瘤相关抗原(TAA)寻找靶点,开发通用治疗型疫苗。
2020年4月,该公司与清大明韵合作,清大明韵将分两批先后注资超过5000万元投入到美诺恒康的mRNA肿瘤疫苗研发项目中。此外,美诺恒康的新冠预防型mRNA肿瘤疫苗正在军事医学科学院开展相关研究。
创始人李林鲜博士,师从Moderna 创始人Robert Langer 教授,成立于2019年,创始人在mRNA特别是LNP递送技术领域拥有多年临床前药物研发经验。
该公司目前在罕见病、肿瘤治疗性疫苗、肿瘤免疫治疗增强剂、感染性疾病预防疫苗四大方向构建研发管线,部分已经进入临床前研究。
深信生物曾入选《麻省理工科技评论》2020年度TR50,上榜理由为“公司与国际mRNA药物研发三巨头在研发方法、递送路径等方面保持同步,独有知识产权的新一代脂质纳米粒(LNP)递送技术处于国际领先水平”。
创始人彭育才,曾任美国 Biogen 公司高级科学家,成立于2019年,在2017年便开始专注于mRNA技术研发,2020年1月获股权融资,投资方为天优投资、横琴金投、康橙投资及丽珠集团。
丽凡达生物在药物设计、生产和制剂递送方面申请了多项发明专利,前期积累了不少疫苗开发经验,产品管线包括传染病疫苗、肿瘤疫苗、罕见病、及其他蛋白缺陷类疾病等,目前进展最快的mRNA狂犬疫苗正处于临床前研究阶段。此外,该公司mRNA新冠病毒疫苗标准样品检测到了目标抗体的产生,已于今年2月交付国家相关权威单位进行动物试验和药效验证。
成立于2018年,致力于建设纳米递送、mRNA合成、疫苗转化三大平台,目前已累计提交4项发明专利和1项实用新型专利。
产品管线上,厚存纳米以LLLRNA纳米递送技术为核心,主要有HPV疫苗、流感疫苗、KRAS疫苗、狂犬疫苗及新冠疫苗五条研发管线。目前与上海细胞治疗集团签署合作基本完成mRNA纳米递送研发平台的建设,与中科院深圳先进院等共同开发新型冠状病毒的mRNA疫苗。
成立于2019年,是国内少数拥有mRNA合成技术和mRNA药物递送系统相关自主知识产权的新药研发公司。
目前瑞吉生正在开发9种预防性mRNA疫苗,进行7项Ⅰ期研究。
成立于 2019 年 6 月落地杭州,创始人王子豪博士在国外药企任职长达11年,期间直接参与过多种疫苗的 CMC 开发,其中 7 年专注于 mRNA 技术平台的工艺开发;联合创始人郭志军领导参与过多种疫苗在中国的临床申报或注册上市。
主要致力于 mRNA 创新药物的研发。该公司基于自复制 mRNA 的核心平台技术可应用于包括肿瘤免疫治疗、传染病疫苗、罕见病治疗、医学美容等领域,尤其是在肿瘤免疫治疗方面有比较独特的优势。部分产品已进入临床申报阶段。
嘉晨西海陆续和欧林生物(SH688319)合作研发 mRNA 新冠疫苗;与康泰生物(SZ300601)合作研发 mRNA 人用狂犬病疫苗;与天境生物(NASDAQ:IMAB)开展技术合作,研发抗体药物;与君实生物(1877.HK,688180.SH)基于 mRNA 技术平台和其它技术平台合作研发肿瘤、罕见病等疾病项目。
2021 年 2 月,嘉晨西海完成过亿元 A 轮融资,由华创资本领投,浙商创投跟投。
2008 年 3 月在国内注册成立,是美国公司 Sirnaomics 在中国的研发与运营主体,创始人陆阳博士是最早投身 RNAi 领域的产学界专家之一。受益于海内外并行布局,圣诺得以在学术资源、研发进度和供应链管理上获得来自美方的优势,同时又能借力国内政策和资本助推、开辟方兴未艾的中国市场。
圣诺主要有多肽纳米颗粒 (PNP) 及新型 GalNAc RNAi 递送平台。PNP 递送平台允许通过局部或静脉内给药,以毒性低、易制造及触达除肝脏外靶器官的能力的显着优点将 siRNA 及 mRNA 递送至患病细胞,而新型 GalNAcRNAi 递送平台能够实现对肝细胞的高效、特定递送。这两个平台能够敲低两个不同的靶基因以产生协同效应,从而提高对抗疾病的治疗潜力。
管线适应症涵盖了癌症、纤维化疾病、代谢疾病和病毒感染等多种疾病,研究进度最快的是其核心候选产品 STP705,已进入 II 期临床研究阶段。
2021 年 7 月,圣诺已经在港交所递交了招股书,拟在香港主板挂牌上市。在此之前,圣诺共完成了 5 轮融资,累计超 2.6 亿美元。
于 2007 年1 月在江苏昆山注册成立,是国内 siRNA 领域的领军企业。
公司已经申请发明专利 60 多项,其中得到授权的有 28 项。
已有包括降血糖、抗前列腺癌、抗视神经损伤、抗乙肝、抗高血脂等多个小核酸药物在内的在研产品管线。该公司目前具有年产公斤级 GalNAc 小核酸原料药的生产能力和分析能力。
公司目前有 4 款产品处于临床研究阶段和临床试验申请阶段,有 10 余款产品处于临床前或早期发现阶段。
其中,SR061 用于治疗非动脉炎性前部缺血性视神经病变 (NAION),处于 II/III 期临床研究阶段,SR062 用于治疗 2 型糖尿病,处于 II 期临床研究阶段,SR063 用于治疗 AR-V7 阳性的转移性去势抵抗性前列腺癌 (mCRPC),处于 IIa 期临床研究阶段,SR016 用于治疗慢性乙肝,处于 IND 申请阶段。
目前,瑞博生物完成了 4 轮融资,累计金额超 10 亿元,资方名单中不乏高瓴、中金等顶级资本,投后估值超 30 亿元。
由李龙承教授在 2016 年创立。RNAa 也是由李龙承于 2006 年在全球首次发现并命名的生物学机制。
不同于 siRNA 或 miRNA 的抑制基因表达机理,由李龙承自研的 RNAa 技术旨在开启内源性基因表达,通过恢复蛋白质的天然功能来治疗疾病。英国公司MiNA Therapeutics 也采用了 RNA 激活技术,已有项目进入临床 I 期阶段。
RNAa 技术能够表达蛋白质的特性,能够解决许多由于蛋白表达不足而致病的情况,弥补现有小核酸疗法只能抑制蛋白表达的空缺。如果产品研发进程顺利,即相当于开辟了全新的细分领域,市场前景十分可期。
中美瑞康建立了 RNA 激活药物发现与开发平台和含有 10 多个新药项目的研发管线,并全部拥有独立自主知识产权,涵盖基因遗传病、肿瘤疾病、靶点在肝脏的代谢及血液系统疾病、皮肤疾病等领域。
2021 年 2 月,中美瑞康完成 1.1 亿元 A 轮融资,由高瓴创投领投,建银医疗成长基金、渤溢资本和隆门资本跟投。
拥有亚洲领先的核酸药开放式技术平台,并在全球范围内提供 CDMO/CMO 整体解决方案。早在 2016 年就获得 NMPA 颁发的寡核酸原料药生产许可证,同时拥有全亚洲最大的 GMP 寡核酸原料药基地。
锐博生物是中国首家且唯一获得 CFDA 药品生产许可证的 cGMP 寡核酸生产基地, 国内第一家做寡核酸药物的 CMO/CRO 公司。公司目前拥有核酸药物生产、核酸分析检测平台、高通量测序、核酸新药研发等八大平台,提供包括RNAi、miRNA、lncRNA、细胞组织分析、生物芯片、高内涵筛选、生物信息学等十大类、40 多种产品及 50 多种服务。
公司创始人张必良博士曾在美国麻省大学医学分子医学系和生物化学与分子药理系任教,主要从事 RNA 化学生物学小干扰 RNA(siRNA)的设计与合成工作。且锐博生物有 2006 年诺贝尔生理/医学奖获得者 CraigC.Mello 坐镇。
2021 年 2 月,张必良汇报了与钟南山团队合作的有关 mRNA 疫苗开发生产的项目,据悉,这款 mRNA 疫苗在 2 到 8℃ 的环境下保存 6 个月仍然有效,这项研发也成为钟南山领衔的攻关项目的子课题,被纳入中国第四批科技部新冠肺炎重点研发计划。
据张必良表示,锐博生物拥有中国最大的 mRNA 原料工厂,每年能生产足够 1 亿人使用的剂量。
1. Malone, R. W., Felgner, P. L. & Verma, I. M. Proc. Natl Acad. Sci. USA 86, 6077–6081 (1989).
2. Malone, R. W. Focus 11, 61–66 (1989).
3. Dimitriadis, G. J. Nature 274, 923–924 (1978).
4. Ostro, M. J., Giacomoni, D., Lavelle, D., Paxton, W. & Dray, S. Nature 274, 921–923 (1978).
5. Melton, D. A. et al. Nucleic Acids Res. 12, 7035–7056 (1984).
6. Krieg, P. A. & Melton, D. A. Nucleic Acids Res. 12, 7057–7070 (1984).
7. Wolff, J. A. et al. Science 247, 1465–1468 (1990).
8. Martinon, F. et al. Eur. J. Immunol. 23, 1719–1722 (1993).
9. Jirikowski, G. F., Sanna, P. P., Maciejewski-Lenoir, D. & Bloom, F. E. Science 255, 996–998 (1992).
10. Conry, R. M. et al. Cancer Res. 55, 1397–1400 (1995).
11. Boczkowski, D., Nair, S. K., Snyder, D. & Gilboa, E. J. Exp. Med. 184, 465–472 (1996).
12. Hoerr, I., Obst, R., Rammensee, H. G. & Jung, G. Eur. J. Immunol. 30, 1–7 (2000).
13. Probst, J. et al. Gene Ther. 14, 1175–1180 (2007).
14. Karikó, K., Kuo, A., Barnathan, E. S. & Langer, D. J. Biochim. Biophys. Acta 1369, 320–334 (1998).
15. Karikó, K., Kuo, A. & Barnathan, E. Gene Ther. 6, 1092–1100 (1999).
16. Karikó, K., Ni, H., Capodici, J., Lamphier, M. & Weissman, D. J. Biol. Chem. 279, 12542–12550 (2004).
17. Karikó, K., Buckstein, M., Ni, H. & Weissman, D. Immunity 23, 165–175 (2005).
18. Warren, L. et al. Cell Stem Cell 7, 618–630 (2010).
19. Aldrich, C. et al. Vaccine 39, 1310–1318 (2021).
20. Kremsner, P. G. et al. Wien. Klin. Wochenschr. https://doi.org/10.1007/s00508-021-01922-y (2021).
21. Jeffs, L. B. et al. Pharm. Res. 22, 362–372 (2005).
22. Geall, A. J. et al. Proc. Natl Acad. Sci. USA 109, 14604–14609 (2012)
撰稿人 | 生命科学前沿
责任编辑 | 邵丽竹
审核人 | 何发















评论
加载更多